

摘 要:奶牛乳房炎每年在全球造成巨大的經(jīng)濟損失,抗生素之前一直被用作治療乳房炎的主要手段。然而,抗生素濫用導致了耐藥菌株的出現(xiàn)和畜產(chǎn)品藥物殘留等問題。隨著國家對食品安全、生物安全等問題愈發(fā)關(guān)注,飼料減抗、禁抗等政策相繼出臺,在奶牛乳房炎防治方面亟需尋求綠色安全的替抗手段。本文對微生態(tài)制劑在奶牛乳房炎防治方面的研究進展進行了綜述,以期為治療奶牛乳房炎和提高乳品質(zhì)量安全提供綠色的替抗思路。
關(guān)鍵詞:乳房炎;微生態(tài)制劑;腸道微生物;免疫;抑菌
奶牛乳房炎主要是由病原微生物侵入引起的奶牛乳腺組織炎性疾病,機械因素、環(huán)境因素、用藥不當?shù)榷伎梢鹉膛H榉垦?/span>[1]。當病原體突破乳頭的防御時,它會進入乳腺并向乳池移動,此時介導身體炎癥反應的主要免疫細胞——巨噬細胞會首先接觸并識別入侵的病原體,然后產(chǎn)生大量趨化因子和一些炎性細胞因子,并進一步誘導大量中性粒細胞在受感染的乳腺中積聚清除病原菌,同時釋放大量炎癥介質(zhì)和ROS,從而導致乳腺發(fā)生炎癥反應(圖1),即乳房炎[2]。奶牛乳房炎臨床檢出較多的致病菌主要有無乳鏈球菌、停乳鏈球菌、金黃色葡萄球菌和大腸桿菌,且這四種常見病原菌均產(chǎn)生了不同程度的耐藥性[3~5];這會導致常規(guī)防治手段愈發(fā)難以見效,將來可能會面臨無藥可用的情況。
美國國家乳房炎委員會根據(jù)乳房及乳汁有無肉眼變化,將乳房炎分為非臨床型乳房炎(即隱性乳房炎)和臨床型乳房炎。隱性乳房炎是奶牛發(fā)病率最高的乳房炎,發(fā)病率是臨床型乳房炎的20~40倍,其會導致產(chǎn)奶量緩慢降低、乳品質(zhì)不斷下降,給奶牛業(yè)造成巨大的經(jīng)濟損失[6]。在奶業(yè)發(fā)達的美國,奶牛養(yǎng)殖場每年的損失中有70%~80%是由隱性乳房炎引起產(chǎn)奶量下降造成的[1],這一情況在我國可能更加嚴峻。
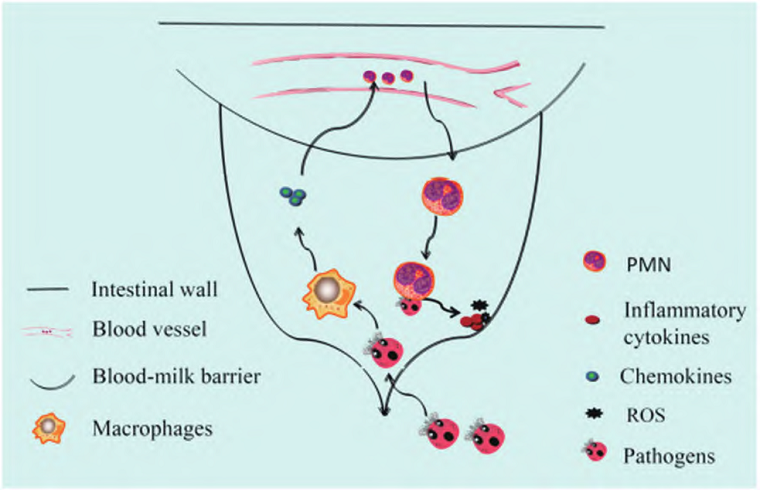
圖1 病原體引起奶牛房炎的發(fā)展路線圖[2]
1 微生態(tài)制劑對奶牛乳房炎的防治作用
臨床上奶牛乳房炎的防治主要依賴抗生素,抗生素的濫用使牧場細菌出現(xiàn)嚴重耐藥性,還造成乳制品中抗生素大量殘留,嚴重威脅人類健康和環(huán)境安全[4]。許多研究表明,微生態(tài)制劑對奶牛乳房炎能起到很好的防治效果。最早,Greene等評估了乳酸菌的乳房內(nèi)輸注是否可以治愈患有亞臨床型乳房炎的奶牛,結(jié)果表明乳酸菌治療的治愈率為21.7%[7],相較于用抗生素頭孢阿司匹林治療的治愈率73.7%存在較大差距,但這讓后面的研究者發(fā)現(xiàn)了益生菌在治療奶牛乳房炎上的潛力。隨后學者們相繼報道了微生態(tài)制劑在降低乳房炎奶牛牛乳中體細胞數(shù)[8~12]、調(diào)控乳汁[12]和腸道[13]微生物上的優(yōu)異表現(xiàn),并發(fā)現(xiàn)其能顯著提高乳品質(zhì)[11,12]和產(chǎn)奶量[9]。楊靜等發(fā)現(xiàn)通過飼喂或乳房灌注微生態(tài)活菌制劑均能對奶牛隱性乳房炎有一定的防治作用,且灌注見效快于飼喂[14]。
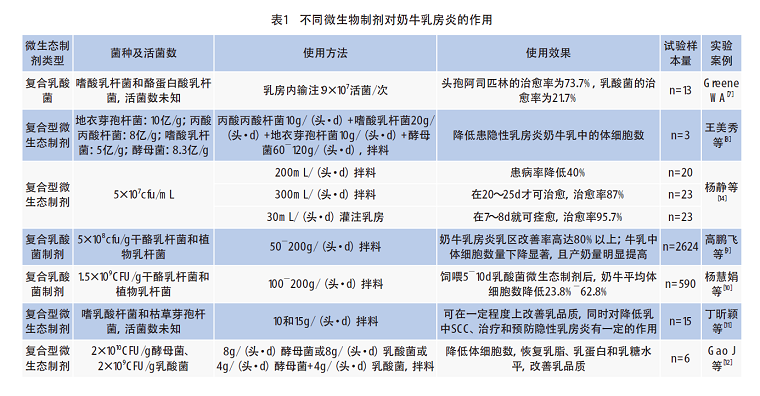
微生態(tài)制劑在實際使用中會受到菌種、活菌含量、使用方法和試驗樣本量等諸多因素的影響,本文將微生態(tài)制劑在奶牛乳房炎上的一些試驗案例從菌種、活菌數(shù)、使用方法、樣本量和實際效果等方面進行了歸納(表1),以供奶牛乳房炎微生態(tài)制劑的研發(fā)及現(xiàn)場使用時借鑒。如表1所示:王美秀等發(fā)現(xiàn)復合型微生態(tài)制劑可降低患隱性乳房炎奶牛乳中的體細胞數(shù)[8],雖然試驗樣本量較少,但也能展現(xiàn)微生態(tài)制劑對乳房炎的有效性;高鵬飛等使用復合乳酸菌制劑對不同地區(qū)牧場的2 624頭奶牛進行試驗,結(jié)果表明復合乳酸菌制劑具有改善奶牛隱性乳房炎、降低牛乳中體細胞數(shù)以及提高奶產(chǎn)量等作用[9];與此同時,楊慧娟等也對不同地區(qū)牧場的590頭奶牛利用復合乳酸菌制劑進行防治試驗,飼喂5~10d后,奶牛平均體細胞數(shù)顯著降低23.8%~62.8%[10]。丁昕穎等利用嗜乳酸桿菌和枯草芽孢桿菌為主要菌種配制的微生態(tài)制劑可在一定程度上改善乳品質(zhì),能夠有效降低乳中SCC[11]。Gao等也發(fā)現(xiàn)日糧中補充乳酸菌和酵母菌能改善乳品質(zhì),顯著降低牛乳中體細胞數(shù),恢復乳脂、乳蛋白和乳糖的含量,且乳酸菌對緩解乳房炎和改變?nèi)橹⑸锏男Ч糜诮湍妇?/span>[12]。
2 微生態(tài)制劑在防治奶牛乳房炎上的作用機理
2.1 調(diào)節(jié)胃腸道菌群及代謝產(chǎn)物
乳房炎奶牛瘤胃中與炎癥相關(guān)的微生物群和代謝物的豐度發(fā)生了顯著變化[12]。腸道微生物群及其代謝產(chǎn)物,包括脂多糖(LPS)和短鏈脂肪酸(SCFAs),已被證明參與了乳房炎的發(fā)展[2]。Ma等通過定量PCR,比較了體細胞數(shù)低(<3×105個細胞/mL)和高(>1×106個細胞/mL)的奶牛的牛奶和糞便微生物特征,觀察到在乳酸菌屬和乳桿菌種水平上存在顯著差異;乳房炎奶牛乳房炎病原體(腸球菌、鏈球菌和葡萄球菌)普遍增加,乳酸桿菌屬(如L. salivarius , L. sakei , L. ruminis ,L. delbrueckii , L. buchneri , L. acidophilus )匱乏;糞便中乳酸桿菌與牛奶微生物群落和SCC相關(guān)聯(lián)[15, 16];隨后他們通過糞菌移植實驗發(fā)現(xiàn)移植乳房炎患牛的糞便能夠引起小鼠乳房炎癥,益生菌干預能夠降低小鼠乳房炎癥程度[13];LPS是革蘭氏陰性細菌細胞壁的主要成分,瘤胃中LPS過度會破壞瘤胃上皮,導致LPS進入血液和肝臟功能受損,LPS經(jīng)血液循環(huán)進入乳腺,增加血液屏障的通透性,導致乳房炎癥;而腸道微生物群產(chǎn)生的SCFAs對乳房炎癥反應有保護作用,并有助于維持血乳屏障的功能[2]。這些結(jié)果表明益生菌能通過調(diào)節(jié)腸道菌群和代謝產(chǎn)物從而對乳房炎有一定的防治作用。
2.2 免疫調(diào)節(jié)
Beecher等將乳酸乳球菌DPC 3147的活菌注入健康牛乳腺,結(jié)果促進了乳汁和血液中IL-1β和IL-8基因表達,刺激宿主發(fā)生免疫反應[17]。Pellegrino等通過微生物學分析表明,在干奶期奶牛乳房內(nèi)接種乳酸菌能增加血液和牛奶中能夠識別金黃色葡萄球菌表位的IgG抗體[18]。日糧中添加乳酸菌和酵母菌能顯著降低乳汁中炎性因子TNF-α、IL-6和IL-1β濃度和髓過氧化物酶(MPO)及N-乙酰-β-d-氨基葡萄糖苷酶(NAG)活性,KEGG分析發(fā)現(xiàn)乳酸菌處理組乳酸菌富集在與乳房炎和炎癥有關(guān)的免疫系統(tǒng)疾病的傳染病通路和免疫系統(tǒng)通路上[12]。鼠李糖乳桿菌能通過促進TLR2和NOD1的協(xié)同作用和減弱不依賴于ASC的NLRP3炎癥小體激活,改善大腸桿菌引起的細胞形態(tài)和超微結(jié)構(gòu)的破壞,限制有害的炎癥反應[19];Li等的研究證明鼠李糖乳桿菌GR-1能誘導PINK1/Parkin介導線粒體自噬,清除受損線粒體并減少ROS 產(chǎn)生和NLRP3 炎性小體激活,減少大腸桿菌誘導的細胞凋亡,并能降低IL-1β、TNF-α等細胞因子水平;從而對大腸桿菌誘發(fā)的乳房炎具有預防和保護作用[20]。
以上這些研究均有力地證明了益生菌能夠增強患乳房炎機體的免疫力,但值得注意的是,不同益生菌菌株對牛乳房炎的免疫調(diào)節(jié)可能存在較大的差異。例如,Fukuyama等發(fā)現(xiàn)嗜酸乳桿菌CRL2074調(diào)節(jié)效果較好,它降低了IL-1α、IL-1β、MCP-1、IL-8和CXCL3的表達,而鼠李糖乳桿菌CRL2084減少了IL-1β、MCP-1和IL-8表達[21];這是由于兩株菌對LPS誘導引起的Toll樣受體負調(diào)控因子調(diào)控的差異性所導致。CRL2074菌株預刺激BME細胞導致Toll樣受體的三個負調(diào)節(jié)因子的表達上調(diào),包括泛素編輯酶A20(也稱為腫瘤壞死因子α誘導蛋白3,TNFAIP3)、單免疫球蛋白IL-1單受體(SIGIRR)和Toll相互作用蛋白(Tollip),而CRL2084預刺激僅上調(diào)Tollip表達[21]。
2.3 抵抗病原微生物
益生菌通過產(chǎn)生細菌素和短鏈脂肪酸[22]、修復黏膜屏障[23,24]和競爭性抑制[25,26]等方式來發(fā)揮抑菌抗炎作用的機制已被廣泛認可。為了解決奶牛乳房炎的問題,研究人員篩選到許多能夠抑制奶牛乳房炎病原菌尤其是金黃色葡萄球菌的益生菌[27~29];Gao等對乳汁微生物測序分析發(fā)現(xiàn),乳酸菌能夠抑制引起乳房炎的腸球菌和鏈球菌繁殖[12]。對于大腸桿菌引起的奶牛乳房炎,Wu等證明鼠李糖乳桿菌GR-1能夠減少大腸桿菌對牛乳腺管細胞(BMECs)的黏附[19]。
3 抗奶牛乳房炎微生態(tài)制劑的研發(fā)
為設計一款綠色安全無殘留、無耐藥性的治療乳房炎的產(chǎn)品,科研人員一直在不遺余力地探索。Espeche等為了設計一種預防牛乳房炎的益生菌產(chǎn)品,從不同奶牛場的初乳樣品中分離出了117株乳酸菌(LAB),并評估了它們的益生性,例如抑制物質(zhì)的產(chǎn)生、表面特性和胞外多糖(EPS)的產(chǎn)生;最終篩到了9株抑菌性能較佳的乳酸菌;并對其進行了抗生素耐藥性和毒力因子等安全性評估[27]。Pellegrino等對牛奶中分離的12株乳酸菌的抗菌活性、對乳房炎致病菌的共聚集及自動抑制情況、對牛乳腺管上皮細胞(BTCEC)的黏附和生長動力學曲線進行了評估,篩選到Lactococcus lactissubsp. lactis CRL 1655和Lactobacillus perolens CRL 1724兩株獨特的乳酸菌,能夠用于預防奶牛干奶期乳房炎的益生菌制劑中[28] 。陳宏偉等從鮮奶和乳制品樣品中篩選獲得5 株能夠抑制金黃色葡萄球菌生長的乳酸菌,并從菌株上清液中提取到細菌素,經(jīng)檢測其具有抑菌活性,抑菌效價分別為457IU/mL、1 023IU/mL、676IU/mL、1 862IU/mL和1 023IU/mL[29]。
抗奶牛乳房炎微生態(tài)制劑的研發(fā)不僅僅限于飼料添加劑。在擠奶前后對奶頭進行消毒是控制奶牛群乳房炎的關(guān)鍵一環(huán),目前該消毒通常用的是化學消毒劑,會存在殘留風險[27];研究人員為了解決這一問題,另辟蹊徑,嘗試用乳酸菌乳頭噴霧劑來替代化藥消毒劑。Alawneh等研究表明乳酸菌噴霧劑與碘噴霧劑對奶牛乳汁體細胞數(shù)的影響遵循類似的趨勢,且與碘噴霧劑處理的奶牛相比,用乳酸菌產(chǎn)品處理的奶牛乳頭末端評分和平均體細胞數(shù)增加的幾率更低[30]。
4 展望
目前,飼料端禁抗、養(yǎng)殖端減抗等政策一步步落地實施,2021年10月21日,為切實加強獸用抗菌藥綜合治理,農(nóng)業(yè)農(nóng)村部農(nóng)牧發(fā)(2021)31號文件制定了《全國獸用抗菌藥使用減量化行動方案(2021—2025年)》,這一政策將極大限制奶牛乳房炎抗菌素用藥量,致使奶牛場的生產(chǎn)特別是針對奶牛乳房炎的替抗已迫在眉睫。微生態(tài)制劑在奶牛乳房炎防治上具有巨大的潛力,無疑是替代抗生素防治奶牛乳房炎的有效選擇之一。對于微生態(tài)添加劑生產(chǎn)企業(yè)來說,如何研制出更能角逐抗奶牛乳房炎的微生態(tài)制劑愈加重要。
作者:李筱雯、黃俊程、喬同、劉爽、周祖濤、肖運才
參考文獻
[1] 郝景鋒. 吉林省奶牛隱性乳房炎主要致病菌敏感中藥篩選與初步應用[D].長春:吉林大學,2018.
[2] Hu X, Li S, Fu Y, et al. Targeting gut microbiota as a possible therapy for mastitis[J]. Eur J Clin Microbiol Infect Dis. 2019;38(8):1409-1423.
[3] 趙高乾, 王素英, 李廣善, 等.奶牛乳房炎主要病原菌感染流行規(guī)律研究進展[J].中國畜牧獸醫(yī), 2019,46(11):3378-3386.
[4] 張行, 李新圃, 嚴勇, 等. 我國奶牛主要養(yǎng)殖區(qū)乳房炎病原菌區(qū)系分布及對抗生素敏感性的最新調(diào)查研究[J]. 中獸醫(yī)醫(yī)藥雜志, 2020,39(04):5-9.
[5] 李宏勝, 羅金印, 李新圃, 等. 我國奶牛乳房炎病原菌區(qū)系調(diào)查及抗生素耐藥性檢測[A]. 中國畜牧獸醫(yī)學會家畜內(nèi)科學分會論文集[C].2011:8.
[6] 高潮. 奶牛隱性乳房炎病原微生物區(qū)系分析及金黃色葡萄球菌基因分型[D]. 合肥:合肥工業(yè)大學,2013.
[7] Greene WA, Gano AM, Smith KL, et al. Comparison of probiotic and antibiotic intramammary therapy of cattle with elevated somatic cell counts[J]. J Dairy Sci, 1991,74(9):2976-2981.
[8] 王美秀, 張愛榮, 郝永清. 防治奶牛隱性乳房炎微生態(tài)制劑的研究[J]. 黑龍江畜牧獸醫(yī), 2008,01:92-94.
[9] 高鵬飛, 程斌, 張善亭, 等. 復合乳酸菌制劑在防治奶牛乳房炎方面的應用研究[J]. 中國畜牧雜志, 2014,50(12):41-47.
[10] 楊慧娟, 張善亭, 崔景麗, 等. 乳酸菌微生態(tài)制劑防治奶牛隱性乳房炎應用研究[J]. 中國奶牛, 2014,17:51-54.
[11] 丁昕穎, 佟桂芝, 張淑芬, 等. 微生態(tài)制劑對奶牛乳成分和體細胞數(shù)的影響[J]. 飼料研究, 2016,13:28-30+44.
[12] Gao J, Liu YC, Wang Y, et al. Impact of yeast and lactic acid bacteria on mastitis and milk microbiota composition of dairy cows[J]. AMB Express, 2020,10(1):22.
[13] Ma C , Sun Z , Zeng B , et al . Cow-to-mouse fecal transplantations suggest intestinal microbiome as one cause of mastitis[J]. Microbiome, 2018,6(1):200.
[14] 楊靜, 高淑琴, 王慶奎, 等. 微生態(tài)活菌制劑防治奶牛隱性乳房炎的效果試驗[J]. 畜牧與飼料科學, 2012,33(08):105-106.
[15] Wang Y, Nan X, Zhao Y, et al. Rumen microbiome structure and metabolites activity in dairy cows with clinical and subclinical mastitis[J]. J Anim Sci Biotechnol, 2021,12(1):36.
[16] Ma C, Zhao J, Xi X, et al. Bovine mastitis may be associated with the deprivation of gut Lactobacillus[J]. Benef Microbes, 2016,7(1):95-102.
[17] Beecher C, Daly M, Berry DP, et al. Administration of a live culture of Lactococcus lactis DPC 3147 into the bovine mammary gland stimulates the local host immune response, particularly IL-1beta and IL-8 gene expression[J]. J Dairy Res, 2009,76(3):340-348.
[18] Pellegrino M, Berardo N, Giraudo J, et al. Bovine mastitis prevention: humoral and cellular response of dairy cows inoculated with lactic acid bacteria at the dry-off period[J]. Benef Microbes, 2017,8(4):589-596.
[19] Wu Q, Liu MC, Yang J, et al. Lactobacillus rhamnosus GR-1 Ameliorates Escherichia coli-Induced Inflammation and Cell Damage via Attenuation of ASC-Independent NLRP3 Inflammasome Activation[J]. Appl Environ Microbiol, 2015,82(4):1173-1182.
[20] Li Y, Zhu Y, Chu B, et al. Lactobacillus rhamnosus GR-1 Prevents Escherichia coli-Induced Apoptosis Through PINK1/Parkin-Mediated Mitophagy in Bovine Mastitis[J]. Front Immunol, 2021,12:715098.
[21] Fukuyama K, Islam MA, Takagi M, et al. Evaluation of the Immunomodulatory Ability of Lactic Acid Bacteria Isolated from Feedlot Cattle Against Mastitis Using a Bovine Mammary Epithelial Cells In Vitro Assay[J]. Pathogens, 2020,9(7):574.
[22] Sanders ME. Impact of probiotics on colonizing microbiota of the gut[J]. J Clin Gastroenterol, 2011,45 Suppl:S115-S119.
[23] Wu H, Xie S, Miao J, et al. Lactobacillus reuteri maintains intestinal epithelial regeneration and repairs damaged intestinal mucosa[J]. Gut Microbes, 2020,11(4):997-1014.
[24] Hou Q, Ye L, Liu H, et al. Lactobacillus accelerates ISCs regeneration to protect the integrity of intestinal mucosa through activation of STAT3 signaling pathway induced by LPLs secretion of IL-22[J]. Cell Death Differ, 2018,25(9):1657-1670.
[25] Kamiya S, Yonezawa H, Osaki T. Role of Probiotics in Eradication Therapy for Helicobacter pylori Infection[J]. Adv Exp Med Biol, 2019,1149:243-255.
[26] Mu?oz-Quezada S, Bermudez-Brito M, Chenoll E, et al. Competitive inhibition of three novel bacteria isolated from faeces of breast milk-fed infants against selected enteropathogens[J]. Br J Nutr, 2013,109 Suppl 2:S63-S69.
[27] Espeche MC, Pellegrino M, Frola I, et al. Lactic acid bacteria from raw milk as potentially beneficial strains to prevent bovine mastitis[J]. Anaerobe, 2012,18(1):103-109.
[28] Pellegrino MS , Frola ID , Natanael B , et al . In Vitro Characterization of Lactic Acid Bacteria Isolated from Bovine Milk as Potential Probiotic Strains to Prevent Bovine Mastitis[J].Probiotics Antimicrob Proteins, 2019,11(1):74-84.
[29] 陳宏偉, 姜云, 郭雪峰, 等. 抑制奶牛乳房炎源金黃色葡萄球菌的乳酸菌的篩選[J]. 中國預防獸醫(yī)學報, 2020,42(02):128-132.
[30] Alawneh JI , James AS , Phillips N, et al . Efficacy of a Lactobacillus-Based Teat Spray on Udder Health in Lactating Dairy Cows[J].Front Vet Sci, 2020,7:584436.
上一篇:沒有了

